
Bitkilerin korkulu rüyası Ralstonia solanacearum hastalığı

Türkçe isimleri: Patates Kahverengi Çürüklük , Domates ve Sardunya Bakteriyel Solgunluk, Muz Moko Hastalığı, Tütn Granville Solgunluğu
İngilizce isimleri: Brown Rot of Potato, Southern Bacterila Wilt of Tomato and Geranium, Moko Disease of Banana, Granville Wilt Of Tobacco
Uluslararası yaygın isimleri:
- English: bacterial wilt; bacterial wilt of solanaceous crops; bacterial wilt of teak; brown rot of potato; brown rot of solanaceous crops; granville wilt of tobacco; moko disease: banana; seedling rot; slime disease: potato; southern bacterial blight of tomato
- Spanish: marchitez bacteriana; marchitez del platano; marchitez del tomate; moco del platano; podredumbre parda de la patata; vaquita de la papa
- French: bactériose vasculaire; bactériose vasculaire de la pomme de terre; flétrissement bactérien de la pomme de terre; flétrissement bactérien de la pomme de terre; flétrissement bactérien de la tomate; flétrissement bactérien des solanacées; flétrissement bactérien du bananier; flétrissement bactérien du tabac; maladie de moko du bananier; pourriture brune
Yöresel yaygın isimleri:
- Germany: Bakterienwelke; Braunfäule der Kartoffel; Schleimkrankheit
Bilimsel ismi:
Ralstonia solanacearum (Smith 1896) Yabuuchi et al. 1996
Sinonim bilimsel isimleri:
- Bacillus musae Rorer 1911
- Bacillus musarum Zeman 1921
- Bacillus nicotianae Uyeda 1904
- Bacillus sesami Malkoff 1906
- Bacillus solanacearum Smith 1896
- Bacterium solanacearum (Smith) Chester 1897
- Bacterium solanacearum var. asiatica (Smith) Magrou 1937
- Bacterium solanacearum var. asiaticum Smith 1914
- Burkholderia solanacearum (Smith 1896) Yabuuchi et al. 1992
- Chromobacterium nicotianae (Uyeda) Krasil’nikov 1949
- Erwinia nicotianae (Uyeda) Bergey et al. 1923
- Erwinia solanacearum (Smith) Holland 1920
- Phytobacterium solanacearum (Smith) Patel & Kulkarni 1951
- Phytomonas ricini Archibald 1927
- Phytomonas solanacearum (Smith) Bergey et al. 1923
- Phytomonas solanacearum var. asiatica (Smith) Stapp 1928
- Pseudomonas batatae Cheng & Faan 1962
- Pseudomonas ricini (Archibald) Robbs 1954
- Pseudomonas solanacearum (Smith 1896) Smith 1914
- Pseudomonas solanacearum var. asiatica (Smith) 1928
- Pseudomonas tectonae Roldan & Andres 1953
- Xanthomonas solanacearum (Smith) Dowson 1943
- Xanthomonas solanacearum var. asiatica (Smith) Elliott 1951
Taksonomisi:
Domain: Bacteria
Phylum: Proteobacteria
Class: Betaproteobacteria
Order: Burkholderiales
Family: Ralstoniaceae
Genus: Ralstonia
Species: Ralstonia solanacearum
Hastalık hakkında genel bilgi :
Ralstonia solanacearum tropik, subtropik ve ılıman bölgelerde yetişen pek çok monokotiledon ve dikotiledon bitki türünün üretimini sınırlayan önemli bakteriyel patojenlerden biridir. Etmenin oluşturduğu hastalık, patateste kahverengi çürüklük, domateste bakteriyel solgunluk, muzda Moko hastalığı gibi farklı isimlerle anılmaktadır. Bakteriyel solgunluk hastalığı ilk olarak 19. yüzyılın sonuna doğru Asya, Güney ABD ve Güney Amerika’ da patates, domates ve yer fıstığında rapor edilmiştir (OEPP/EPPO, 2004). Bakteri ilk defa Smith (1896) tarafından Bacillus solanacearum olarak isimlendirilmiştir. Daha sonraki yıllarda etmenin içerisinde farklı ırk, biovar, RFLP gruplan, phylotipe ve sequevar belirlenmiştir.
Dünyada ki zarar raporları:
Ralstonia solanacearum görüldüğü yerlerde önemli ürün kayıplarına neden olabilmektedir. En büyük ekonomik kayıp Güneydoğu Amerika, Endonezya, Brezilya, Kolombiya ve Güney Afrika’da patates, tütün ve domates bitkilerinde rapor edilmiştir (EPPO/CABI, 1997). Filipinler’de 1966-1968 yıllarında domateste ortalama %15, patlıcan ve biberde %10 ve tütünde %2-5 kayıp bildirilmiştir (Zehr, 1969). Peru’da Amazon havzasının muz plantasyonlarının yaklaşık yansında hastalık görülmüş ve hızlı yayılış göstererek Peru ormanları boyunca plantasyonlan yok olma riski ile karşı karşıya bırakmıştır (French and Sequeira, 1968).
Hindistan’da domateste bazı yıllarda ürün tamamen yok olmuştur (EPPO/ CABI, 1997). Yunanistan’da patateste 1951-1953 yıllarında önemli kayıplar oluşmuştur (Zachos, 1957). İsrail’de ise ürün kayıplan ilkbahar üretimi patateslerde, sonbahar üretimi patateslere göre daha ağır olmuştur (Volcani and Palti, 1960). Hastalık şiddeti etmenin kök nematodlan ile birlikte bulunması durumunda artmaktadır (Sitaramaiah and Sinha, 1984).
Patates, domates, muz, tütün dışında etmenin diğer önemli konukçulan arasında Anthurium spp., biber (Capsicum annuum), pamuk (Gossypium hirsutum), patlıcan (Solanum melongena), zencefil (Zingiber officinalis), sardunya (Pelargonium zonale), dut (Morus spp.) bitkileri ve Solanum nigrum, Solanum dulcamara, Portulaca oleracea gibi bazı yabancı otlar yer almaktadır (Hayward, 1994 a; EPPO/CABI, 1997; OEPP/EPPO, 2004).
Ülkemizde ki zarar raporları:
Ralstonia solanacearum ilk önce 1995 yılında Nevşehir’de beş patates tarlasında saptanmış olup buradan eradike edilmiştir (Özakman ve ark., 1998). Daha sonra 2006 yılında Ege Bölgesi Balıkesir ili, Ayvalık ilçesi, Altınova beldesi yemeklik patates alanlarında etmenin varlığı belirlenmiştir (Ustun et al., 2008). Aynı yıl Ege Bölgesi Çanakkale ili Batak ovasında ise etmen sofralık domateslerde, sulamada kullanılan bir kanal suyunda ve Solanum nigrum, Solanum alatum ve Portulaca oleracea gibi yabancı otlarda bulunmuştur (Ustun et al., 2007a; Ustun et al., 2007b).
Nasıl bir bakteridir ?
Değişik konukçularda solgunluk hastalığına neden olan etmen Ralstonia solanacearum (Smith) Yabuuchi et al. ’dır. Etmenin sinonimleri; Bacterium solanacearum (Smith) Chester, Burkholderia solanacearum (Smith) Yabuuchi et al. (1992) ve Pseudomonas solanacearum (Smith) Smith’dir. Ralstonia solanacearum infraspesifık düzeyde değişkenlik gösteren bir etmendir. Farklı ırkları, biovarlan, biotipleri, fllogrupları bulunmaktadır.
Patojenisite özelliklerine göre 5 ırkı bilinmektedir (Buddenhagen, 1962; Buddenhagen 1986).
Irk-1 bütün dünyada tropik bölgelerde görülmekte ve tütün, diğer solanaseler ve farklı familyalardan pek çok bitki türünü hastalandırmaktadır. Aynca bu ırk, ırk 2, 4 ve 5’te olduğu gibi yüksek sıcaklık optimuma (35°C’de) sahiptir.
Irk 2 genelde Güney Amerika’nın tropik bölgelerinde mevcut olup muz ve Heliconia bitkilerinde ‘’Moko” olarak adlandınlan hastalığa neden olmakta, aynı zamanda Filipinlerde muzlarda “Bugtok” denilen hastalığı oluşturmaktadır.
Irk 3 (patates ırkı) tropik, subtropik ve ılıman bölgelerin yüksek yerlerinde patates başta olmak üzere domates, sardunya (Pelargonium zonale), patlıcan, biber ve Solanum nigrum ve Solanum dulcamara gibi bazı yabancı ot solanase türlerini etkilemektedir (OEPP/EPPO, 2004). Aynı zamanda ırk 3 solanacea olmayan yabancı ot türlerinde de asimptomatik olarak bulunabilmektedir (Pradhanang et al., 2000; Janse et al., 2004). Bu ırk düşük sıcaklık optimuma sahip (27°C) olup 35°C’nin üzerinde ve de 10°C’nin altında virulensliği azalmaktadır (Stansbury et al., 2001).
Asya’da görülen ırk 4 zencefile (Zingiber ojfîcinalc),
Çin’de bulunan ırk 5 ise dut (Morus spp.)’a özelleşmiştir.
Biyolojik özellikleri
Etmenin içinde karbonhidrat ve alkollerin kulanımındaki farklılıklara göre 5 farklı biovar (biotip) belirlemiştir (Hayward 1964; Buddenhagen 1986). Etmenin ırklan ve biovarlan arasında ilişki saptanmamıştır (Buddenhagen, 1962). Sadece ırk 3’ün biovar 2 ile uyuştuğu bildirilmiştir. Biovar 2’nin içerisinde 3 ayrı subphenotype belirlenmiştir.
Birbirinden trehalose, D-ribose ve meso-inositol kullanımı bakımından farklılık gösteren bu fenotipler Cook et al. (1989)’un belirlediği bazı RFLP gruplan ile uyumlu olup Biovar 2 A (RFLP grubu 26), 2A (RFLP grubu 27) ve Biovar 2T veya 2N olarak bilinmektedir. Dünyada en yaygın olan ve genelde ılıman ve subtropikal bölgelerde veya tropik bölgelerin yüksek yerlerinde bulunan Biovar 2 A (RFLP grubu 26) D-ribose negatif, trehalose negatif ve meso- inositol pozitiftir. Biovar 2 A (RFLP 27) ise D- ribose negatif, trehalose pozitiftir ve mesoinositol negatif olup sadece Şili ve Kolombiya (Güney Amerika)’nın bazı bölgelerinde bulunmaktadır.
Biovar 2 N veya 2 T (RFLP 29-31) D-ribose pozitif, trehalose pozitif ve meso-inositol pozitiftir ve genelde Peru ve Brezilya’da görülmektedir. RFLP (Restriction Fragment Length Polymorphism) analizi ile etmenin içerisinde 46 multilocus genotip (MLG) belirlenmiş ve bu MLG’ler divisyon 1 (“Asiaticum”) ve divisyon 2 (“Americanum”) olarak adlandırılan 2 ana strain kümesinde birleştirilmiştir (Cook et al. 1989; Cook, and Sequeira, 1994). Irk 1 (biovar 3, 4, 5)’in Asya strainleri birinci grubu oluştururken ırki (biovar 1), ırk 2 (biovar 1) ve ırk 3 (biovar 2)’ün Amerikan strainleri ikinci grubu oluşturmuştur.
16S-23S arasında bulunan bölgedeki farklılıkları baz alarak yapılan phylogenetik analiz sonunda da 4 phylotip belirlenmiştir (Fegan and Prior, 2005). Phylotype 1 – genellikle amerikan kaynaklı Biovar 3,4 ve 5 strainleri; Phylotype II dünyanın farklı yerinden izole edilen biovar 1 ırk 1; biovar l ırk 2 (Moko hastalığı strainleri), biovar 2 /ırk 3 ve biovar 2T strainleri; Phylotype III-Afrika kökenli biovar 1 ve 2T strainleri, Phylotype IV- Endonesya, Japonya (bazı strainler) ve Avustralya (tek strain) kaynaklı biovar 1, 2 ve 2T strainlerini kapsamaktadır. Her phylotype, endoglucanase (egt) geninin bir kısmının sekanslamasmdaki farklılıklara dayanarak sequevarlara ayrılmıştır.
Yaşam Döngüsü ve Epidemiyoloji
Bakteri toprakta ve yüzey sularında yaşayabilmektedir (EPPO/CABI, 1997). Bu ortamlarda konukçusu olan yabancı otların varlığında yaşam süresi uzamaktadır. Tropik bölgelerde pek çok yabancı ot türü alternatif konukçuluk ve patojeni bir üründen diğer ürüne taşıyan köprü görevini yapmaktadır.
Bitkiye giriş yaralanmış kök ve gövde veya stomalar yoluyla gerçekleşmektedir. Bitki içerisinde bakteri sistemik olarak iletim demetlerinde hareket etmekte ve solgunluk oluşmaktadır. İletim dokusundaki ilerleme hızı, sıcaklık ve kolonize edilen bitki organı gibi faktörlere bağlı olarak değişmektedir. İletim demetlerinin bakteri tarafından tıkanması solgunluğun ana nedenidir.
Hastalık 24-35°C derecede daha şiddetli seyretmektedir. Kış aylarında, ortalama sıcaklığın 10°C’nin altına düştüğü ılıman bölgelerde nadiren görülmektedir. Farklı ırkların (biovarlann) optimum hastalık gelişimini sağlayacak sıcaklık istekleri değişmektedir (Swanepol, 1990).
Yüksek toprak nemi ve uzun süren nemli veya yağmurlu havalar hastalık şiddetini arttırmaktadır. Toprak nemi aynı zamanda etmenin yaşayışı ve çoğalmasını etkilemektedir (Nesmith and Jenkins, 1985).
Uygunsuz çevre koşullan belirti çıkışını etkilemektedir. Örneğin Kenya’da 1520-2120 m yükseklikte üretilen tohumluk patates yumrulan belirti göstermezken daha düşük yükseklikte üretildiklerinde belirti göstermiş ve bu durum yumruların latent olarak enfekte olmasma bağlanmıştır (Nyangeri et al., 1984).
Etmenin ırklanmn çoğu doğal yolla çok sınırlı yayılmaktadır. Ancak, Muzda Moko hastalığına neden olan ırk 2 böcek vektörlerle taşındığından yüksek doğal yayılım potansiyeline sahiptir. Irk 3 biovar2 enfekteli Solanum dulcamara bitkileri barındıran yüzey sularıyla kolaylıkla yayılabilmektedir. Bulaşık yüzey sularının sulamada kullanılmasıyla bakteri diğer konukçulara yayılmaktadır (Olsson, 1976).
Uzun mesafeli (uluslararası) yayılmada en önemli yol latent olarak enfekteli tohumluk patates yumrulan ve diğer vejetatif çoğaltım materyallerinin kullanılmasıdır (EPPO/CABI, 1997). Doğal olarak tohumla taşınma sadece yer fıstığı için geçerlidir (EPPO/CABI,1997). Irk l’in domates, biber ve patlıcan tohumlarında bulunabilmesi ile ilgili az sayıda çalışma bulunmaktadır (Persley, 1986b; Kelman et al, 1994; Singh, 1995). Uygunsuz çevre koşullarında, kısmi dayanıklı çeşitlerde veya strainlerin virulensliği düşük olduğunda patates yumrularında latent enfeksiyonlar görülebilir (EPPO/CABI.1997). Latent enfeksiyonlar yeni alanların bulaşmasında en büyük etkendir.
Hastalığın Belirtileri
Patates bitkisinde ki belirtiler:
Yeşil akşamda, ilk belirtiler, gündüz sıcaklığın en yüksek olduğu saatlerde dalların uçlarındaki yapraklarda görülen solgunluk şeklindedir. Solgunluk erken enfeksiyonlarda gece serinliğinde kaybolur. Bu dönemde yapraklar yeşil kalır, ancak daha sonra sararma, bronzlaşma ve kahverengi nekrozlaşma gelişir. Kısa süre içerisinde tek taraflı veya tüm bitkide görülen solgunluk, geriye dönüşümsüz hale gelir. Hastalık ilerledikçe kök boğazının hemen üzerindeki bölgede çizgi şeklinde kahverengileşme oluşur, bitki çöker ve ölür.
Hasta bitkinin enine kesitinden iletim dokusunun genelde kahverengileştiği anlaşılır ve kesilen yüzeyden sütümsü beyaz bakteriyel akıntı çıktığı gözlenir. Bu akıntının iplikçik şeklindeki çıkışı, kesilmiş gövde dikey olarak şeffaf cam kap içinde bulunan suya batırıldığında, açıkça görülür. Patatesi hastalandıran diğer bakteriyel etmenler böyle bir akıntı oluşturmaz (EPPO/ CABI,1997). Bu nedenle suda akıntı izleme testi tarlada ön teşhis için önerilir.
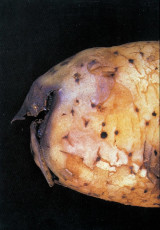
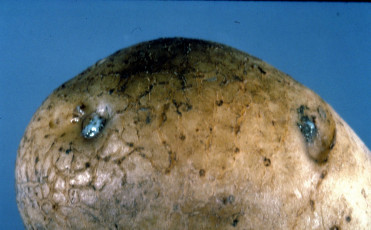
Yumrularda ki belirtileri:
Yumrular göbek bağı (heel end)’na yakın yerden enine veya göbek bağının üzerinden geçecek şekilde boyuna kesildiğinde görülür (EPPO/CABI, 1997). Erken enfeksiyonlarda, halka şeklindeki yumru iletim dokusu cam sarısından açık kahverengine kadar renk değiştirir ve buradan birkaç dakika sonra kendiliğinden açık krem renginde bakteriyel akıntı çıkar. Daha sonra iletim dokusundaki renk değişimi daha belirginleşir, koyulaşır ve parankima dokusunda ilerleyen nekroz görülür.
İleri evrelerde, göbek bağı ve yumrunun diğer gözlerinden yapışkan bakteriyel akıntı dışarıya çıkar ve bu noktalara toprak partikülleri yapışır. Yumru iletim dokusunun çökmesi nedeniyle bazen kabukta kırmızı-kahve renginde hafif çukur lekeler oluşabilir. Hastalığın ileri döneminde fungal ve bakteriyel etmenlerin sekonder gelişmesi sonucunda yumuşak çürüklüğün meydana gelmesi mümkündür.
Yumru belirtileri, Patates halka çürüklüğü hastalığı (Clavibacter michiganensis subsp. sepedonicus) ile karıştırılabilir. Ancak yumru göbek bağı ve gözlerinden çıkan bakteriyel akıntıya toprak par- tiküllerinin yapışması kahverengi çürüklük hastalığı için tipik olup halka çürüklüğünde görülmemektedir. Aynca kahverengi çürüklük hastalığı ile bulaşık yumrular kesildiğinde birkaç dakika içinde iletim demetlerinden kendiliğinden krem renginde bakteriyel akıntı çıkar, halka çürüklüğü hastalığında ise daha sarımsı olan bakteriyel akıntının iletim demetlerinden çıkması için yumrunun sıkılması gerekir (EPPO/CABI, 1997).
Kahverengi çürüklük hastalığının yeşil aksam belirtilerini taşıyan bitkiler hem hasta hem de sağlıklı yumrular oluşturur. Sağlıklı görünen bitkiler ise bazen latent olarak enfekteli yumrular meydana getirir.













Domates bitkisinde ki belirtileri:
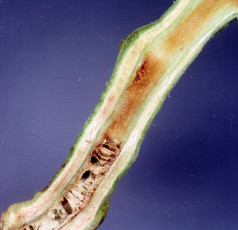
İlk belirtiler, genç yaprakların turgorunu kaybederek sarkmasıdır. Çevresel koşullar uygun olduğunda (toprak sıcaklığının yaklaşık olarak 25°C ve toprağın neme doygun olması), bir kaç gün içerisinde tek taraflı veya tüm bitkiyi saran solgunluk gelişir (EPPO/CABI, 1997). Kısa sürede bitkiler ölür. Solgun bitkilerin gövdeleri boyuna kesildiğinde, iletim demetinin renk değiştirdiği ve buradan kendiliğinden beyaz veya sarımsı bakteriyel akıntının çıktığı görülür.
Gövdenin alt kısımlarındaki öz dokusu kahverengileşir. Bazen gövde üzerinde havai kökler oluşur. Çevresel koşullar hastalık gelişimi için yeterince uygun olmadığında (toprak sıcaklığı 21°C’nin altında) daha az solgunluk oluşur, ancak gövde üzerinde çok sayıda havai kök meydana gelir (Mc Carter, 1991). Tarlada, hastalığın ön teşhisi için patateste yeşil aksam belirtileri bölümünde yer alan suda akıntı izleme testi yapılabilir.














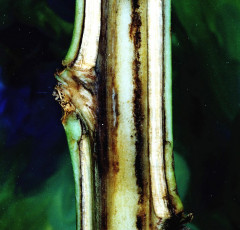
Tütün bitkisinde ki belirtileri:
En önemli belirtilerden biri tek taraflı solgunluk ve zamansız sararmadır. Bitkinin bir tarafında bulunan yapraklarında hatta yaprağın yansında solgunluk olabilir. Şiddetli enfeksiyonlarda, yapraklar sararmadan solar ve gövdeye asılı kalır. Domateste olduğu gibi bitki boyuna kesildiğinde, iletim dokusunun renk değiştirdiği görülür. Birincil ve ikincil kökler kahverengiden siyaha kadar renk değiştirebilir (Echandi, 1991).











Muz bitkisindeki belirtileri:
Ralstonia solanacearum’un oluşturduğu Moko hastalığının belirtileri Fusarium oxysporum f.sp. cubense etmeninin neden olduğu belirtilerle kolayca karışabilir. Ancak meyvede belirti gözlendiğinde bu iki hastalığı ayırmak mümkün olur, çünkü kahverengi ve kuru çürüklük sadece Moko hastalığında görülür. Genç ve hızlı gelişen bitkilerde, yeni gelişen yapraklar açık yeşil veya sarıya döner ve çöker. Bir hafta içinde tüm bitkilerin yapraklan çökebilir, Genç fışkınlarda siyahlaşma, gelişme geriliği veya kıvrılma görülebilir. Yalancı gövdelerde iletim demetleri kahverengileşir (Hayward, 1983).









Sardunya bitkisinde ki belirtileri:
Erken belirtiler alt yapraklarda kıvrılma ve solgunluk, kısmi sararma ve nekroz şeklindedir. Yapraklarda, bazen bakteriyel yanıklık hastalığı (Xanthomonas pelargonii)’ nda olduğu gibi V şekilli lekeler oluşur (Douglas, 2002). Gövdelerde toprağa yakın kısımlarda ve iletim dokusunda kahverengi-siyah renk değişimi görülebilir. Sistemik olarak gelişen solgunluk başlangıçta gece ve serin havada iyileşir, ancak daha sonra kalıcı hale gelip iyileşme olmaz (Douglas, 2002).









Mücadelesi
Karantina önlemleri:
Ralstonia solanacearum ülkemiz dahil EPPO ülkeleri, ABD vb. gibi pek çok ülkenin karantina listesinde yer almaktadır. Bu ülkelerde etmene karşı karantina tedbirleri uygulanmaktadır. Avrupa Birliği ülkeleri, en yüksek yayılma potansiyeline sahip ırk 3/biovar2’nin önlenmesini ve eradikasyonunu esas alan ortaklaşa mücadele direktifi (EU, 1998) hazırlamıştır. Ülkemizde Avrupa Birliği’nin bu mevzuatı doğrultusunda 29 Eylül 2002 tarih ve 24891 sayılı Resmi Gazetede “Ralstonia solanacearum (Smith) Yabuuchi et al.’m Mücadelesi Hakkında Tebliğ” yayınlanmış ve mevzuat hükümlerinde karantina tedbirlerinin nasıl yürütüleceği belirtilmiştir.
Kültürel önlemler
Hastalıksız üretim materyali kullanımı:
Ralstonia solanacearum etmeni vejetatif üretim materyalleri ile taşındığından üretimde hastalıksız sertifikalı üretim materyalleri kullanılmalıdır. Etmenin üretim materyallerinde varlığının saptanması için klasik (yan seçici besiyerine izolasyon, biyokimyasal ve pathojenisite testleri), serolojik (immuno-fluorescence-IF, ELISA) ve moleküler (PCR) yöntemleri birleştiren standart teşhis protokolleri mevcuttur (OEPP/EPPO, 2004).
Ekim nöbeti:
Topraktaki inokulumu yok etmek amacıyla konukçusu olmayan bitki türleri ile en az 5-7 yıllık ekim nöbeti önerilmektedir (EPPO/ ÇABI.1997). Patatesin mısır veya fasulye ile dönüşümlü olarak yetiştirilmesi bazı durumlarda inokulum yoğunluğunu ve hastalık çıkışım azaltmakta (Autrique and Potts, 1987), ancak bu konukçularda etmen, canlılığım sürdürebildiği raikrolesyon oluşumuna neden olmaktadır (Granada and Sequeira, 1983).
Toprak koşullarının değiştirilmesi:
Gübre uygulamalarıyla toprak Ph’sını değiştirmek mücadele yöntemi olabilir. ABD’de, toprak Ph’sını yazın 4-5’e düşürüp sonbaharda 6’ya yükselterek etmen eradike edilmiştir (EPPO/CABI, 1997). Toprağa yapılan bazı ilaveler topraktaki mikrobial aktiviteyi arttırarak inokulumu düşürmektedir. Örneğin toprağa kümes hayvanları gübresi ve çiftlik gübresi ilavesi domateste bakteriyel solgunluk hastalığım azaltmıştır (İslam and Toyota, 2004).
Konukçu yabancı ot mücadelesi:
Etmen çok sayıda yabancı otta kalabildiğinden yeni bulaşmalara kaynaklık eden bu otlarla mücadele çok önemlidir. Sadece tarlalarda bulunan otlar değil aynı zamanda sulama kanallarının kenarlarında yetişen Solanum dulcama gibi bitkiler de yok edilmelidir. Avrupa ülkelerinde etmenin eradikasyonu için su kanallarının kenarında yetişen Solanum dulamara‘ya karşı herbisit uygulaması yapılmış ve başarılı sonuçlar alınmıştır (Persson, 1998)
Kendi gelişen bitkilerle mücadele:
Hastalığa kaynaklık eden ve tarlalarda kendiliğinden yetişen domates, patates, tütün vb. gibi konukçu bitkiler imha edilmelidir.
Sanitasyon:
Hastalığın bulaşmasını önlemek için bu önlemlerin önemi büyüktür. Kesilmiş yumruların tohumluk olarak kullanılması hastalık oranını 2.5 kat arttırmakta ve %>40 ürün kaybına neden olmaktadır (Vijayakumar et al., 1985). Bu nedenle yumrular kesilmeden dikilmelidir. Etmenle bulaşık su kaynaklannın sulamada kesinlikle kullanılmaması gerekmektedir, çünkü yüzey suların etmenin yayılmasını sağlayan en kolay yollardan biridir. Üretimde kullanılan tüm makine, alet ve depolama alanlan temizliğe tabi tutulmalı ve uygun bir dezenfektan ile muamele edilmelidir.
Hastalıkla bulaşık olduğu tespit edilen üretim yerlerinde yetişen bitkisel materyal hastalık riski taşımaktadır. Bu materyalin özel çöp fırınlarında yakılması veya tanmsal alanlara veya sulamada kullanılan kaynaklanna sızmayacak şekilde derin gömülmesi gerekmektedir.
Nematod mücadelesi:
Bitki köklerinde yara açan nematodlarla hastalık arasında sinerjestik etki olduğundan nematodlarla mücadele edilmelidir.
Dayanıklı çeşit:
Ralstonia solanacearum ’un farklı ırk ve biovarlarının bulunması nedeniyle dayanıklı çeşit geliştirme zordur. Bu hastalığa karşı farklı bitki türlerinde geliştirilen dayanıklılık polygenik olup çevresel koşullara bağlılık göstermektedir (Kelman, 1953; Hayward, 1991). Kolombiya’da Solanum phureja ve Solanum demissum kullanılarak yapılan melezlemelerle dayanıklı patates çeşitleri elde edilmiştir (French, 1985; Hartman ve Elphistone, 1994).
Başka çalışmalarda da orta ve yüksek derece dayanıklı klonlar saptanmış ancak bu klonların yumrularında etmenin latent olarak bulunabildiği bildirilmiştir (Priou, 2001; Priou et al., 2005). Bazı yabani genotiplerde (Solanum acaule ve Solanum tuberosum subsp. andigena) yumruda latent enfeksiyon oluşmadan yüksek derecede dayanıklılık bulunmuştur (Priou, 2006). Domateste Ralstonia solanacearum ‘a dayanıklılık yabani Lycopersicum esculentum var cerasiforme ve Lycopersicum pimpinellifoliım kaynaklıdır (Scott et al., 2005). Bu nedenle küçük meyve boyu ile ilişkili olup (Acosta et al., 1964; Wang et al., 1998) büyük meyveli çeşitlere aktarılması güç olmuştur (Scott et al., 2005). Ancak Hawaii 7997 çeşidinden yüksek dayanıklılığa sahip büyük meyveli hatlar elde edilmiştir (Scott et al., 2007).
Kimyasal mücadele:
Hastalığın etkin bir kimyasal mücadelesi bulunmamaktadır. Toprak fumigantların etkisi çok düşük veya hiç yoktur (Murakoshi and Takahashi, 1984). Streptomycin, ampicillin, tetracycline and penicillin gibi antibiyotikler de etkisizdir (Farag et al., 1982). Mısır’da streptomycin uygulamalarının hastalık oranını attırdığı bildirilmiştir (Farag et al., 1986).
Biyolojik mücadele:
Bacillus polymyxa ve Pseudomonas flııorescens antagonistleri laboratuar koşullarında etmeni kontrol etmiştir (Aspiras and Cruz, 1985). Şili’de Pseudomonas fluorescens patateste laboratuarda ve tarlada hastalığa karşı etkili bulunmuştur. Bazı çalışmalarda avirulent mutantlar da kullanılmaktadır (Gallardo & Panno, 1989; Hartman & Elphinstone, 1994).
Yazılarımızdan haberdar olmak için
Abone olmak istermisin ?
Kaliteli yazılarımız ve fotoğraflarımızdan ilk sizin haberiniz olsun !
Bitki koruma ailesine katıldığınız için teşekkür ederiz.
Bir şeyler yanlış gitti sanırım 🙁




